
导读
土壤产甲烷菌参与复杂的相互作用,决定了群落的结构和功能。研究探究了土壤产甲烷菌的共现模式、影响因素以及其对甲烷产量的影响(主要由物种相互作用调节)和这些相互作用的功能意义。本研究测量了亚洲大陆各地稻田中的甲烷排放量,并将共现网络中成对关系表示为产甲烷古菌群落的共现模式中涉及的复杂相互作用。结果表明,共现网络的拓扑学参数与年平均温度呈正相关,是所有生物和非生物因素中甲烷排放的最重要预测因子。39个局域网络中参与共存关系的产甲烷类群对甲烷排放的贡献最大(53.3%),远高于特定链接的产甲烷类群(36.8%)。潜在的关键类群属于Methanobacterium,Methanocella,Methanothrix,Methanosarcin,与甲烷产生功能基因mcrA,fwdB,mtbA和mtbC高度相关。此外,共存的分类群表现出非常不同的构建模式,约30%的确定性和约70%的随机性。相反,更高比例的随机性(93~99%)表征了特定地共存分类群的构建模式。这些结果表明,微生物的共存模式与其功能意义密切相关,普遍共存的潜在重要性进一步揭示了相互作用的复杂网络可能比物种多样性对土壤功能的贡献更大。
论文ID
原名:Coexistence patterns of soil methanogens are closely tiedto methane generation and community assembly in rice paddies
译名:土壤产甲烷菌的共生模式与稻田甲烷的产生和群落聚集密切相关
期刊:Microbiome
IF:11.607
发表时间:2020.01
通讯作者:梁玉婷
通讯作者单位:中国科学院南京土壤研究所
本研究于2013年8月至2013年11月从中国北方至南方13个地区的39个稻田中采集了429个土壤样品(图1a)。稻田代表了四种农作物轮作管理方式(单稻,稻麦轮作,双稻,三联稻)以及五种土壤类型(来源于炉坍黄土的中性黑土,来源于黄河冲积物的碱性潮土,来自湖泊沉积物的水状水稻土,来源于第四纪红土的酸性红土,来自浅海沉积物的淹育水稻土),并收集了13个采样区域的地理信息,包括经度,纬度,年平均温度(MAT)和年平均降水量(MAP)。在每个采样场中,采用空间上“L形”采样设计在100 m×100 m的地块内采集了11个土壤样本。从表土(0–15 cm)中随机选择五个直径为5cm的中心点,每个子采样点的直径为0.5 m,并混合在一起(总计500 g),在实验室中将土壤分为两个子样本。将一个子样品保持在4°C下以测量土壤地化性质,包括pH,壤总有机碳(SOC),溶解有机碳(DOC),总氮(TN),硝酸盐(NO3--N),铵(NH4+-N),总碳(TP),有效磷(AP),总钾(TK),有效钾(AK)以及CH4产生潜力。将另一个子样品提取DNA进行测序分析。
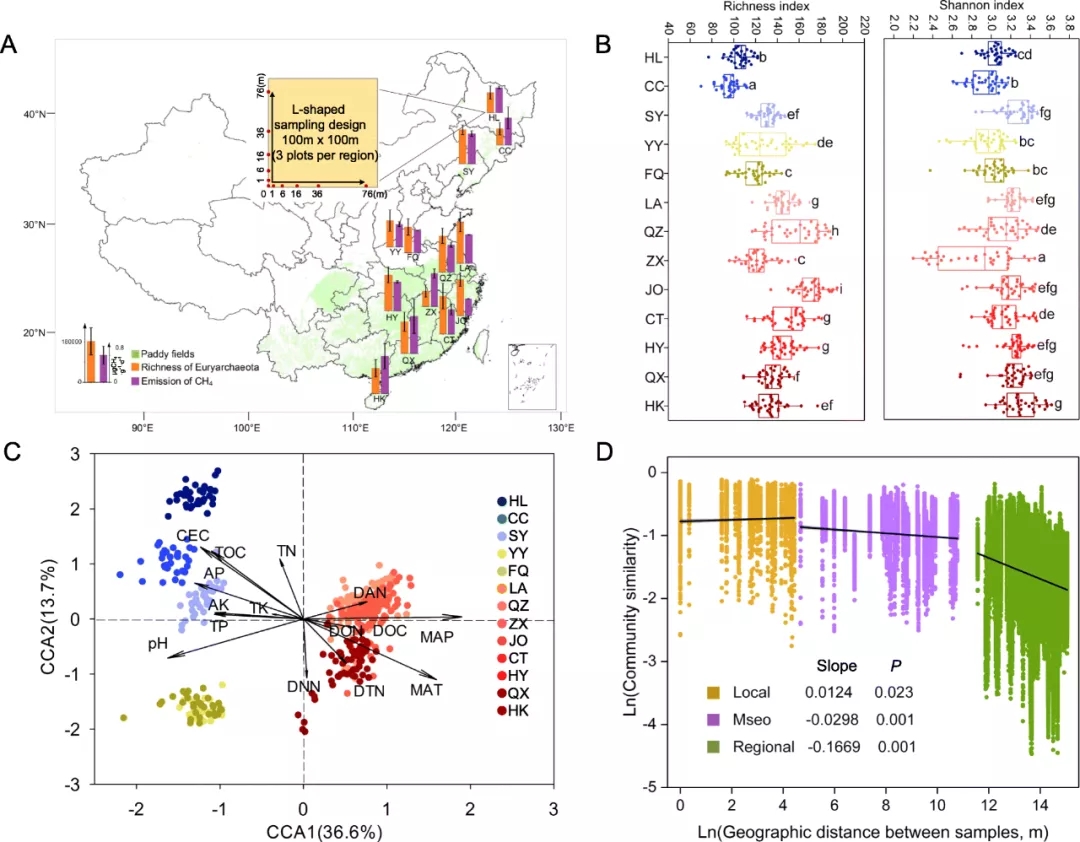
图1 稻田土壤甲烷生成潜力和产甲烷古细菌群落分布。a来自中国北部至南部39个典型稻田(绿色)的产甲烷古细菌群落的丰富度(橙色)和CH4排放潜力(紫色)。b产甲烷菌的α多样性(丰富度和香农指数)。方框内的水平条表示中位数。框的顶部和底部分别代表第75个百分点和第25个百分点。c产甲烷古细菌群落结构的典型对应分析(CCA)。黑色箭头指示解释变量的向量,不同颜色的点指示39个稻田中的稻田土壤样品(总共429个土壤样品)。MAT,年平均温度;MAP,年平均降水量;CEC,阳离子交换能力;TOC,总有机碳;DOC,溶解有机碳;TN,总氮;DTN,溶解的总氮;DON,溶解的有机氮;AN,铵氮;DNN,硝酸盐氮;TP,总磷;AP,有效磷;TK,总钾;AK,可用钾。d三种规模的产甲烷群落的距离-衰减关系,局部(1–100 m),中间(0.1–50 km)和地区(100–3500 km)。
结果
1 稻田土壤产甲烷菌群落的多样性
稻田中的甲烷排放差异很大(图1a)。所有429个样品的多样性稀释曲线表明,测序深度捕获了大多数微生物信息。产甲烷的α-多样性随采样位点而变化(图1b),并且与MAT呈显著正相关(P<0.05,ANOVA)。甲烷排放与土壤产甲烷菌的Shannon-Wiener多样性相关。
产甲烷的群落沿不同气候带的地理位置分别聚类(图1c)。研究发现MAT,MAP,pH,CEC和TOC显著影响产甲烷菌群落分布(P<0.05)。距离衰退关系的定义意味着群的相似性随着地理距离的增加而降低。在局部尺度上,距离衰退关系呈现出正斜率,这意味着在很小的空间尺度(1–100 m)上,产甲烷菌群落不存在距离衰退关系(图1d)。此外,随着规模的增加,群落的相似性降低,线性拟合斜率从-0.0298到-0.1669。这些结果表明产甲烷群落表现出清晰的生物地理分布格局。
2 产甲烷菌群落的共现网络
基于OTU水平构建了所有样品(图2a)和每个局域样品的产甲烷菌群落共现网络。局域网络的架构从北到南逐渐变得复杂化。节点数从39增加到147,边数从45增加到1039。Methanoregula,Methanothrix,Methanocella,Methanosarcina,Methanobacterium(属),Methanomicrobiales和Methanosarcinales(目)占据节点的比例很高,其范围从62.2(QZa)到95.0%(HLb)。
对于主要的环境因子来说,MAT是八个网络拓扑学属性(节点数,边数,正相关,负相关,模块性,平均聚类系数,网络直径,特征路径长度)之间变化的最强预测因子。线性回归分析进一步表明,MAT与所有网络属性均呈显著正相关(P<0.05)(图2c)。此外,我们发现丰富度也对网络结构有影响。通过控制变量,探索了温度和丰富度对网络的重要性,结果表明温度更为重要(R = 0.254> R =-0.004)。以上表明,相比其他环境因素,MAT与产甲烷菌群落的网络结构更紧密相关。
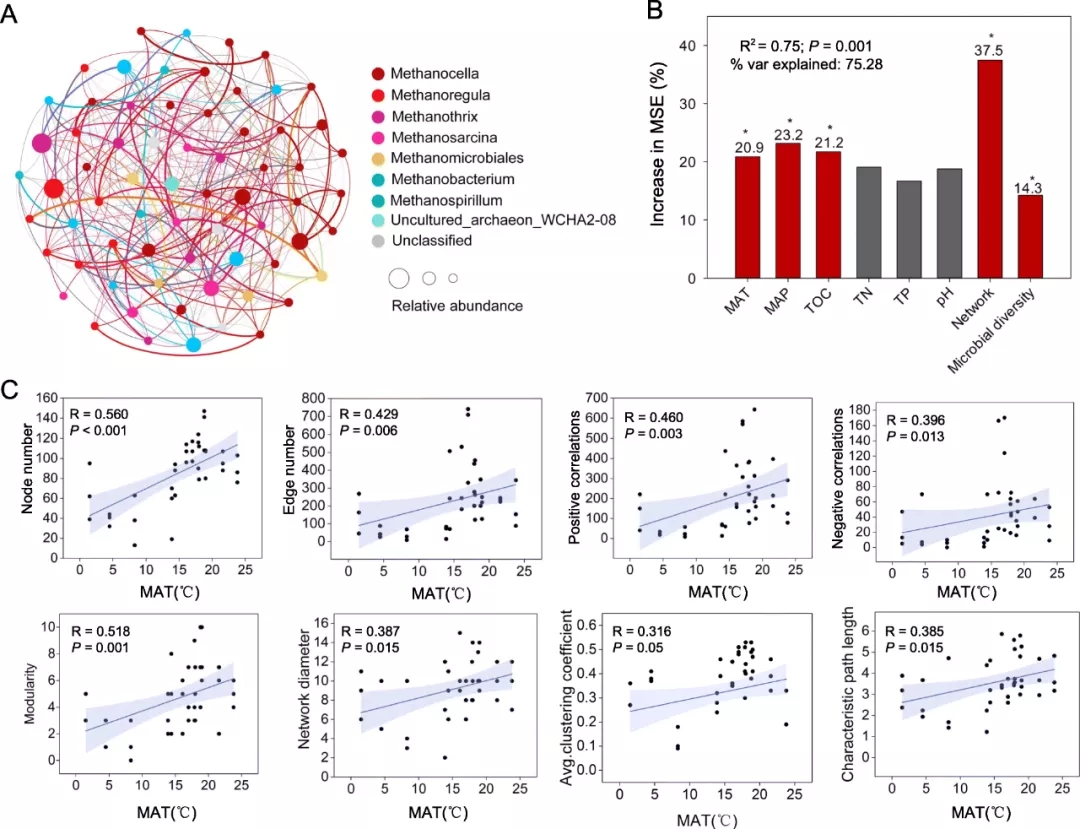
3 土壤产甲烷菌中普遍和特有的共存类群
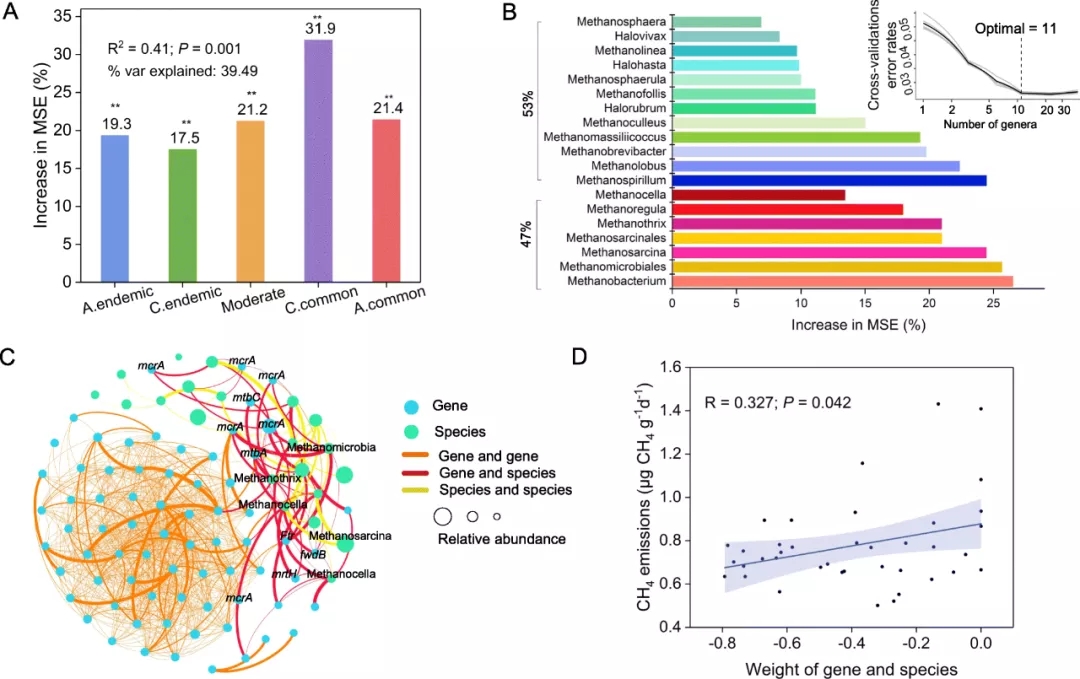
与地化因子(MAT和MAP)和主要土壤因子(TOC,pH,TN和TP)相关的产甲烷群落(共生网络和多样性)因素解释了甲烷排放总量变化的75.28%,其中产甲烷菌之间的相互作用占最大百分比(37.5%)(图2b)。线性回归分析表明,在八个拓扑学参数中,模块性、平均聚类系数和网络直径与甲烷排放呈显著正相关(P<0.05)。以上结果表明,这些产甲烷菌的共同存在可能是甲烷排放的主要因素。
为解决这一假设,即共有和特有同时存在的产甲烷菌在维持群落结构和功能中可能发挥不同的作用,根据这39个网络中边出现的频率,将所有39个网络的成对共现关系分为5组:总是特有,有条件特有,中等,有条件共有和总是共有,相关的OTU数量分别为314、209、131、32和9。五组的微生物丰富度和香农多样性显著不同(P <0.05,ANOVA,Duncan test)。通过构建一个随机森林模型来预测五组共发生关系对甲烷排放的影响(图3a)。共有(有条件共有和总是共有)共发生关系的贡献率最高(53.3%),而特有(有条件特有和总是特有)共发生关系的贡献率为36.8%。因此,通常共存的33个OTU可能被认为是甲烷排放的潜在重点。在33个关键类群和功能基因之间观察到紧密的关系(图3c)。基因与基因、物种与基因、物种与物种的边缘权重之和分别为1.26、1.01和0.40,表明关键类群与功能基因之间存在较强的联系。线性回归分析表明物种和基因相关权重与甲烷排放量呈正相关。
通过GeoChip检测到的33个潜在的关键基因和甲烷生成功能基因之间建立了功能网络(图3c),92个节点共产生1119条边。在关键类群和功能基因之间观察到密切的关系。基因与基因,物种与基因以及物种与物种之间的边缘权重之和分别为1.26、1.01和0.40,这表明关键类群与功能基因之间存在相对较强的联系。参与这些相互作用的基因和种类主要是mcrA,fwdB,mtbA,mtbC,Methanocella,Methanothrix,Methanosarcina,Methanobacterium。线性回归分析表明,物种和基因联系的权重与甲烷排放呈显著正相关(P <0.05)(图3d)。
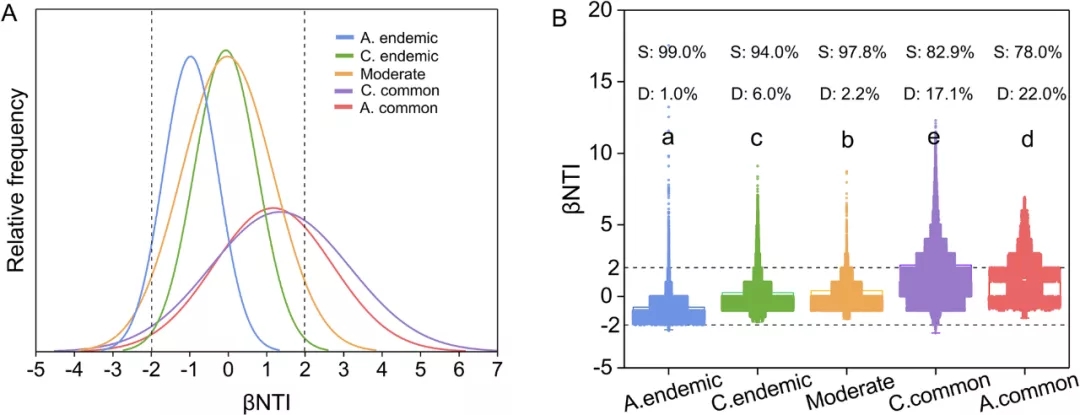
4 普遍和特有的共存类群不同的群落组装方式
五个组的βNTI值主要在−2和2之间:总是特有(98.76%),有条件特有(93.2%),中等(97.58%),有条件共有(74.21%),总是共有(71.52%)。群落组装以随机过程为主,而常见共存分类单元的确定性过程所占比例增加。常见共存类群的βNTI值分布更广泛,且频率相对较低(图4)。其他组的βNTI以相对较高的频率集中在2和-2之间。
讨论
使用网络分析来探索在复杂多样的群落中共存的微生物分类群之间的直接和/或间接作用,可能有助于确定许多微生物群在基本生态学和生活史策略中的功能作用和组装过程。根据本研究的假设,即跨大陆稻田的MAT介导产甲烷菌的复杂共现关系,我们发现网络的所有拓扑属性都与MAT正相关。平均聚类系数和特征路径长度的增加表明,高度连接的OTU在其附近分组并且非随机地聚在一起。
一种解释是温度会长期影响稻田中产甲烷菌的独特群落组成。根据生态学和纬度多样性梯度,温度升高土壤微生物丰富度和空间异质性的代谢理论,参与产甲烷水稻田土壤乙酸和丙酸转化的古细菌和细菌群落结构均随梯度温度从25°C开始变化到50°C。拓扑学参数变化的另一个潜在解释是环境过滤会影响微生物竞争和共生。适应一种微生物的环境压力可能会增加/降低另一种微生物的选择压力,从而产生拮抗/交感合作相互作用。我们的结果表明,MAT与产甲烷菌的正向相互作用具有更高的相关系数,而不是负向相互作用。温度可能增强产甲烷古菌的协同作用,这对稻田的群落功能具有重要影响。在本研究中,与微生物多样性相比,共存关系可以更好地预测甲烷排放的变化(图2b)。该结果与先前基于网络的研究一致,即当网络连接指数从0.626增加到1.278时,土壤食物网对碳的吸收从50%增加到75%,这表明网络结构与生态系统功能过程紧密相关。分类单元之间的紧密联系可能有助于揭示群落成员共有的潜在生态位占用特征。此外,长期的地质过程赋予当代环境持久的遗产。地质过程直接或间接影响生物多样性和生态系统功能。例如,先前的研究表明,气候变化的区域尺度变化可以确定生物多样性对自然生态系统中生态系统多功能性的影响。Hu等人探索了生物群落的主要驱动力,发现MAT对细菌群落的影响最大。另外,地质过程可能导致生物形态和演化。Poltak等人提出了一种进化方案,其中古细菌的祖先具有甲烷代谢能力。物种之间的共同进化可以增强生态系统特征;例如,物种进化出互补的资源利用方式,从而提高了生态系统的生产力。
在共生模式的研究中,总体趋势(通才边)和局部信号(专家边)在适应环境因素以及对抗/交感合作互动中的共存模式中的作用至关重要。本研究中超过72%的边被确定为特有边(39个网络中边数量≤3的频率),其中超过75%的OTU处于产甲烷群落的共现关系中。然而,特有边对甲烷生成的贡献(36.8%)远低于普遍边(53.3%)。这个结果与专家比通才消耗资源更快的观点相矛盾。
根据使用情况(即栖息地通才和栖息地专家),我们进一步寻求对潜在关键类群进行分类。共同联系中涉及的潜在关键物种包括Methanosarcinaceae,Methanocellales,Methanobacteriales,Methanomicrobiales。这些分类单元在维持群落中重要的公共关系方面可能具有很强的适应性。例如,Methanosarcinaceae也可以使用H2/CO2作为底物,尽管效果不如Methanobacteriaceae和Methanocellaceae。Methanosarcinaceae包括acetoclastic methanogens和其他通用产甲烷菌,Methanocellaceae,Methanomicrobiacea,Methanoregulaceae和Methanobacteriaceae都是氢营养型的。由产乙酸细菌形成的乙酸盐既可以被某些产甲烷菌(Methanosarcina spp和Methanosaeta spp)直接使用,也可以被细菌的同养结合(例如,同养乙酸酯氧化剂)和耗氢的产甲烷古生菌降解。氢营养型产甲烷作用可以通过种间电子转移充当电子的汇,从而减少形成氢的产乙酸细菌和耗氢古细菌之间的当量。在先前的研究中,氢营养型产甲烷菌可以维持在低的溶解H2浓度下并产生乙酸盐。温度影响微生物群落的整体多样性。先前的研究证实,在不同温度下,不同种类的产甲烷菌可能会占优势。最著名的产甲烷菌是嗜温的和中等至极度嗜热古细菌。例如,Methanobacteriaceae在30°C时表现出最大的活性,而Methanocellaceae在45°C的后期表现出最大的活性。对于乙酸发酵型甲烷化产甲烷菌,Methanobacteriacea可以在中等温度(10–30°C)下通过氢营养和乙酸发酵型甲烷化过程产生甲烷,而在较高温度(45°C)下仅消耗H2/CO2而不是乙酸盐,它们的丰度在8°C–21.6°C的范围内最高。与文献中报道的最佳生长温度相比,它们并不完全相同。我们认为,这种灵活的策略引起了生态系统过程和功能的转变,从而提高了甲烷的生产效率。
我们还发现,关键分群与参与产甲烷过程的功能基因mcrA,fwdB,mtbA和mtbC高度相关(图3c)。甲基甲酰胺:CoM甲基转移酶(mtbA)在甲基营养型甲烷生成过程中参与将CO2还原成甲烷和乙酸盐歧化成甲烷和CO2的过程。这些过程可能是关键物种主导甲烷排放的原因。这项研究涉及的功能基因都与甲烷的产生有关(21个功能基因),在先前的研究中已经进行了广泛的研究和验证,例如mcrA,fwdB和mtbA。在本研究中,GeoChip芯片用于测量包括甲烷代谢过程在内的功能基因的丰度。因此,由于GeoChip是无法发现新功能基因的闭环系统(除非已发现新基因并将其放入GeoChip库中),因此可以通过网络分析来识别更多可用基因。因此,我们基于网络分析筛选出的所有功能和关键基因,并进行了统计和分析确认。我们希望在将来的研究中找出更多与产甲烷菌的结构和功能有关的基因。
与产甲烷相关的群落组装过程同时受到确定性和随机过程的影响。共生物种通过在环境过滤和扩散限制之间进行平衡来适应环境条件,从而改变其他物种的选择压力以及它们如何利用可用资源。物种选择是确定性过程,其定义为由于生物之间的适应度差异和环境异质性而改变群落结构的生态力。相反,分散限制可以是确定性的,随机的或两者兼有。焦等人指出,物种选择导致在低纬度稻田中古细菌的共现关联往往更为频繁。由于在本研究中MAT是主要的环境过滤器,我们发现随着共现关系频率的增加,βNTI的分布向边缘移动(βNTI=2)。确定性过程对群落组装的相对贡献具有相似的趋势,并伴随着共现关系的增加。尽管随机过程在驱动微生物群落的组装中仍起着主导作用,但我们的结果表明,经历强大的环境选择(MAT)和生存适应的共同存在的分类单元更可能相互关联,并在甲烷排放中起着更重要的作用。先前的一项研究表明,生物选择(物种选择)比水稻土中的其他作用对微生物的组装过程的贡献更大,这一推论可以得到证实。由于产甲烷作用受到不同的温度过滤作用,共同存在的分类单元之间更紧密的共存关系会出现,生态位占有率更高,从而提高了甲烷的生产效率。在未来的研究中,可以考虑将这种共存关系添加到气候变暖模型(例如GISS全球气候模型)中,以提高模型预测的准确性。
此外,水稻在不同发育阶段对产甲烷古细菌群落组成的影响将影响甲烷排放。Kimura等指出,稻根向根际供应的各种化合物的类型和数量随生长期的不同而变化。与以前的研究一致,一些关键类群,如Methanosarcinaceae,Methanobacteriales,和Methanomicrobiales,在水稻生长期中会显著影响稻田甲烷的排放。因此,尽管在本研究中我们没有考虑农作物对产甲烷菌组成和功能的影响,但农作物的作用不容忽视。在未来的研究中,我们需要研究这些关键物种对水稻生长不同阶段甲烷排放的作用,并分析水稻生长对这些微生物群落,结构和功能的影响。
评论本文研究了大陆范围内对稻田中产甲烷菌的共现模式,发现了MAT高度介导了紧密的网络结构。在产甲烷的微生物功能中普遍共存关系可能比特定共存更重要。随机过程和确定性过程的相对重要性在普遍共存的群落和特定性共存的群落之间是不同的。这些结果表明,微生物共存模式与群落的功能意义密切相关,特别是普遍共存的分类群的重要性,进一步表明复杂的相互作用网络可能比物种多样性对土壤功能的贡献更大。需要进行野外和实验室实验来进一步研究产甲烷菌共存模式,该模式在生态时间尺度上驱动群落组成和功能以及物种相互作用的进化。